Искусственный интеллект, нейросети
193.7 K
453
1.2 K


Малая Вишера
Карма: -21.94
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Цитата: slavae от 09.11.2017 17:52:00Отсюда

абсолютно предсказанный результат - автомат в открытом мире недееспособен
"Мне плевать на вас ублюдки.
Я анархо-аморал." (С)
Я анархо-аморал." (С)
- +0.13 / 10
-

Карма: -2.99
Регистрация: 13.08.2015
Сообщений: 3,725
Читатели: 8
Аккаунт заблокирован
Регистрация: 13.08.2015
Сообщений: 3,725
Читатели: 8
Аккаунт заблокирован
Искусственный интеллект, к сожалению - наша единственная надежда. Глядя, с каким наслаждением Хомо избавляется от приставки Сапиенс, очень хочется верить в то, что удастся создать хоть какой-то бэкап, для перезагрузки системы.
Технические средства вероятно уже превысили возможности мозга, они не востребованы для развития и направляются в основном на поддержание комфортного состояния. Например, одно время казалось, что Всемирная Сеть Интернет(тм), позволив соединить потенциал людей, находящихся вне физического контакта, приведёт к возникновению нового качества и уровня мышления. Это было смешное заблуждение.
Видимо, социум исчерпал возможности развития. Остаётся надеяться на то, что искусственная среда справится с решением задач оценки и выработки решений более эффективно, оставив человечеству право на комфортное потреблебление.
Технические средства вероятно уже превысили возможности мозга, они не востребованы для развития и направляются в основном на поддержание комфортного состояния. Например, одно время казалось, что Всемирная Сеть Интернет(тм), позволив соединить потенциал людей, находящихся вне физического контакта, приведёт к возникновению нового качества и уровня мышления. Это было смешное заблуждение.
Видимо, социум исчерпал возможности развития. Остаётся надеяться на то, что искусственная среда справится с решением задач оценки и выработки решений более эффективно, оставив человечеству право на комфортное потреблебление.
- -0.03 / 7
-
Малая Вишера
Карма: -21.94
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Цитата: AndreyK-AV от 11.11.2017 23:10:13Работа, это кроме добывание средств, ещё и самоутверждение, самоуважение и амбиции, необходимые для сохранения потребности в разуме у общества двуногих, прямо ходящих млекопитающих.

проявления животной сущности человека, вызванной половым влечением и как следствие амбиции вами перечисленные не имеют никакого отношения к разуму
"Мне плевать на вас ублюдки.
Я анархо-аморал." (С)
Я анархо-аморал." (С)
- +0.04 / 4
-

Москва
49 лет
Карма: +2.33
Регистрация: 07.03.2008
Сообщений: 277
Читатели: 0
Регистрация: 07.03.2008
Сообщений: 277
Читатели: 0
Цитата: slavae от 13.07.2017 11:44:55Отсюда
Google's DeepMind AI just taught itself to walk
Google's artificial intelligence company, DeepMind, has developed an AI that has managed to learn how to walk, run, jump, and climb without any prior guidanc...
Перевод текста из видео:
Выглядит, конечно, смешно...
...но данная анимация показывает что–то совершенно восхитительное.
Это Искусственный Интеллект DeepMind от Google учится ходить.
Все движения, что делают эти забавные фигурки...
...ИИ пришлось изобрести самому.
Модель получила виртуальные сенсоры,
Которые сообщают собственную ориентацию в пространстве
И расположение объектов вокруг.
Затем программисты стимулируют ИИ двигаться из точки А в точку B.
И все.
ИИ не рассказывали, что такое "ходить",
Это то, до чего ему пришлось додуматься самому.
Модель может бегать по пересеченной местности,
Прыгать,
Карабкаться и переваливать через препятствия,
И держать равновесие когда двигаешься вперед.
Получается не всегда.
Но на ошибках учатся.
ИИ тренировался на трех разных моделях.
У четырехногой в итоге особенно хорошо получились прыжки.
А гуманоид выработал довольно необычную походку.
Возможно он знает что–то, чего не знаем мы.
Да, ИИ сегодня делает "первые шаги". Нам они кажутся смешными и неуклюжими, как у годовалого ребенка.
Только учится он быстрей. И побежит дальше чем мы.
Слава роботам!
Вот еще: https://player.vimeo.com/video/79098420
Это не ИИ, а просто генетический алгоритм для обучения ходьбе произвольных существ. Например, как научить бегать жирафоподобного динозавра. На мой взгляд, гораздо реалистичнее получилось. Если бы это скрестить с ИИ выше, получилось бы идеально.
- +0.03 / 4
-

Карма: +39.26
Регистрация: 05.06.2010
Сообщений: 20,434
Читатели: 8
Регистрация: 05.06.2010
Сообщений: 20,434
Читатели: 8
Цитата: l-mik от 13.11.2017 06:18:01Вот еще: https://player.vimeo.com/video/79098420
Это не ИИ, а просто генетический алгоритм для обучения ходьбе произвольных существ. Например, как научить бегать жирафоподобного динозавра. На мой взгляд, гораздо реалистичнее получилось. Если бы это скрестить с ИИ выше, получилось бы идеально.
ИИ ценен тем что приобретенный им опыт может легко тиражироваться и передаваться другим ИИ-особям, но для этого опыт должен приобретаться в реальных обстоятельствах. Опыт приобретенный в виртуальной реальности имеет смысл лишь как отработка соответствующего софта.
- +0.00 / 0
-
Москва
49 лет
Карма: +2.33
Регистрация: 07.03.2008
Сообщений: 277
Читатели: 0
Регистрация: 07.03.2008
Сообщений: 277
Читатели: 0
Цитата: Поверонов от 13.11.2017 09:16:03ИИ ценен тем что приобретенный им опыт может легко тиражироваться и передаваться другим ИИ-особям, но для этого опыт должен приобретаться в реальных обстоятельствах. Опыт приобретенный в виртуальной реальности имеет смысл лишь как отработка соответствующего софта.
Для роботов виртуальный макет очень близок к реальному, поэтому умение двигаться и держать равновесие можно передать другим экземплярам с такой-же конструкцией. И отточить эти навыки на виртуальной модели. Вот в этом ролике проходит 900 итераций, пока не научится. Будем 900 экземпляров делать?
- +0.01 / 1
-
Карма: +39.26
Регистрация: 05.06.2010
Сообщений: 20,434
Читатели: 8
Регистрация: 05.06.2010
Сообщений: 20,434
Читатели: 8
Цитата: l-mik от 13.11.2017 11:02:05Для роботов виртуальный макет очень близок к реальному, поэтому умение двигаться и держать равновесие можно передать другим экземплярам с такой-же конструкцией. И отточить эти навыки на виртуальной модели. Вот в этом ролике проходит 900 итераций, пока не научится. Будем 900 экземпляров делать?
О том и речь. В данном случае идет отработка конструкции самого робота для удержания равновесия. Примитивный ИИ здесь лишь для проверки управляемости конструкции корпуса робота.
- +0.00 / 0
-
Малая Вишера
Карма: -21.94
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
поскольку мы находимся в динамическом мире масштабируемом силой времени единственная задача мозга - расчёт координат точек пространства в будущем
Знания позволяют нам предсказывать большее количество координат пространства в будущем в квант времени - знания это коэффициент ускорения в бесконечно большое будущее.
При этом мозг не оперирует никакими численными методами - только последовательное сравнение больше/меньше. Для принятия решения требуется либо до 30% для ответа нет и 70% и более для ответа да.
Создать в общем-то не проблема. Вопрос как заставить это работать без внешней силы человека. Цель человеческого мозга вычисление/предсказание всех координат бесконечного пространства - стимулом вычислений является смерть.
Знания позволяют нам предсказывать большее количество координат пространства в будущем в квант времени - знания это коэффициент ускорения в бесконечно большое будущее.
При этом мозг не оперирует никакими численными методами - только последовательное сравнение больше/меньше. Для принятия решения требуется либо до 30% для ответа нет и 70% и более для ответа да.
Создать в общем-то не проблема. Вопрос как заставить это работать без внешней силы человека. Цель человеческого мозга вычисление/предсказание всех координат бесконечного пространства - стимулом вычислений является смерть.
"Мне плевать на вас ублюдки.
Я анархо-аморал." (С)
Я анархо-аморал." (С)
- +0.00 / 0
-

47 лет
Карма: +3.89
Регистрация: 11.08.2010
Сообщений: 4,060
Читатели: 0
Регистрация: 11.08.2010
Сообщений: 4,060
Читатели: 0
Цитата: mark.76 от 15.11.2017 09:58:17//////
стимулом вычислений является смерть.
смерть стимулом являться не может .
стимулом в живом мире являются боль и удовольствие , доступные даже одноклеточным .
эти примитивные стимулы задают стимулы более высокого уровня - страх боли и предвкушение удовольствия .
смерть , особенно насильственная , ассоциируется с исключительной болью , поэтому страх боли индуцируется в страх смерти .
а нежелание этот страх испытывать в инстинкт самосохранения .
"что-бы получить правильный ответ надо задать правильный вопрос." - народная мудрость .
"что-бы задать правильный вопрос надо знать большую часть ответа." - Роберт Шекли "ответчик".
"что-бы задать правильный вопрос надо знать большую часть ответа." - Роберт Шекли "ответчик".
- +0.04 / 3
-
Карма: +4.92
Регистрация: 01.04.2011
Сообщений: 1,430
Читатели: 0
Регистрация: 01.04.2011
Сообщений: 1,430
Читатели: 0
Цитата: VladimirKox от 05.11.2017 17:30:22В начале октября 2017 в Москве прошла конференция "Нейроинформатика 2017" http://neuroinfo.ru/…?year=2017 . Где можно ознакомиться с материалами трудов?
Подсказали где можно посмотреть видео с 17 докладов и лекций https://vimeo.com/channels/neuroinfo2017 , т.е. - не секретно. Странная ситуация, со всей страны собираются спецы, что-то там обсуждают, и - концы в воду.
У меня доклад будет в середине декабря в ЮНЦ РАН, ни сослаться на материалы трудов, ни похвалить...
- +0.00 / 0
-
Карма: +0.02
Регистрация: 23.11.2017
Сообщений: 1
Читатели: 0
Регистрация: 23.11.2017
Сообщений: 1
Читатели: 0
Цитата: vlad21 от 05.04.2017 01:17:40Технофобы – люди, боящиеся роботов, искусственного интеллекта и новых технологий, которые они не понимают – скорее всего, опасаются потерять работу из-за этих современных технологий...
 ну и зря.
ну и зря.Их и нанимать обратно на работу скоро будет тоже искусственный интеллект: Инновационное будущее рекрутинга
Удачи всем,
Пётр
- +0.02 / 1
-
Карма: 0.00
Регистрация: 19.12.2017
Сообщений: 1
Читатели: 0
Регистрация: 19.12.2017
Сообщений: 1
Читатели: 0
57 лет
Карма: +7.60
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Цитата: __Alex_loki_ от 16.11.2017 07:54:52смерть стимулом являться не может .
стимулом в живом мире являются боль и удовольствие , доступные даже одноклеточным .
эти примитивные стимулы задают стимулы более высокого уровня - страх боли и предвкушение удовольствия .
смерть , особенно насильственная , ассоциируется с исключительной болью , поэтому страх боли индуцируется в страх смерти .
а нежелание этот страх испытывать в инстинкт самосохранения .
Надуманно... ЗЫ в курсах що рыбы (таки позвоночные - то есть на достаточно высоко ступени развития) не испытывают боль.
- +0.00 / 0
-
57 лет
Карма: +7.60
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Цитата: l-mik от 13.11.2017 11:02:05Для роботов виртуальный макет очень близок к реальному, поэтому умение двигаться и держать равновесие можно передать другим экземплярам с такой-же конструкцией. И отточить эти навыки на виртуальной модели. Вот в этом ролике проходит 900 итераций, пока не научится. Будем 900 экземпляров делать?
Вот это такое настоящий интеллект
...Это не простейшие дискретные классификаторы -нейросети, на тренинг которых затрачиваются десятки человеколет и мегаватты мощности, и на которые теперь по неизвестным причинам стало молиться половина планеты (причем один основных арггументов адептов - это ведь круто потому шо - никто не знает как оно работает ............ыыыыыы) - Новый тотем TensorFlow
PS
Канзи можно оставить в стороне, хотя он и гений, но его учили...А вот с воронами и попугаями весьма поучительно...Ждем когда первый робот с нейросетю такого же размера додумается до подобных достижений в отношении собственной подзарядки.
- +0.38 / 2
-

Москва
Карма: +194.19
Регистрация: 21.03.2013
Сообщений: 27,983
Читатели: 7
Регистрация: 21.03.2013
Сообщений: 27,983
Читатели: 7
Цитата: stranger1234 от 20.12.2017 00:15:26Вот это такое настоящий интеллект
Это не простейшие дискретные классификаторы -нейросети, на тренинг которых затрачиваются десятки человеколет и мегаватты мощности, и на которые теперь по неизвестным причинам стало молиться половина планеты (причем один основных арггументов адептов - это ведь круто потому шо - никто не знает как оно работает ............ыыыыыы) - Новый тотем TensorFlow
PS
Канзи можно оставить в стороне, хотя он и гений, но его учили...А вот с воронами и попугаями весьма поучительно...Ждем когда первый робот с нейросетю такого же размера додумается до подобных достижений в отношении собственной подзарядки.
Тут было видео, как нейросеть научилась передвигаться, имея только цель и возможность шевелить конечностями.
Думаю, она научилась бы и подзаряжаться, будь у неё вилка, а в виртуальном пространстве — розетка.
Империя - это мир, и этой идеологии достаточно. Мы живём в самой лучшей стране в мире и все нам завидуют.
Одушевлённое Одевают, Неодушевлённое Надевают.
Одушевлённое Одевают, Неодушевлённое Надевают.
- -0.08 / 1
-
Малая Вишера
Карма: -21.94
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Цитата: slavae от 20.12.2017 05:49:12Тут было видео, как нейросеть научилась передвигаться, имея только цель и возможность шевелить конечностями.
Думаю, она научилась бы и подзаряжаться, будь у неё вилка, а в виртуальном пространстве — розетка.
Когда?
пример с вороной лишь подтверждает тот факт, что без передачи далее, знания не имеют смысла
"Мне плевать на вас ублюдки.
Я анархо-аморал." (С)
Я анархо-аморал." (С)
- +0.00 / 0
-
Москва
Карма: +194.19
Регистрация: 21.03.2013
Сообщений: 27,983
Читатели: 7
Регистрация: 21.03.2013
Сообщений: 27,983
Читатели: 7
Цитата: mark.76 от 20.12.2017 06:00:08Когда?
пример с вороной лишь подтверждает тот факт, что без передачи далее, знания не имеют смысла
Тут всего 6 страниц )
на 4-й
https://glav.su/forum/2/3061/messages/4473645/#message4473645
Империя - это мир, и этой идеологии достаточно. Мы живём в самой лучшей стране в мире и все нам завидуют.
Одушевлённое Одевают, Неодушевлённое Надевают.
Одушевлённое Одевают, Неодушевлённое Надевают.
- +0.00 / 0
-
Карма: +4.92
Регистрация: 01.04.2011
Сообщений: 1,430
Читатели: 0
Регистрация: 01.04.2011
Сообщений: 1,430
Читатели: 0
Цитата: mark.76 от 20.12.2017 06:00:08без передачи далее, знания не имеют смысла
Это классика! В гранит и бронзу!
Внесу и я. свою лепту
В.С. КОХАНОВ
E-mail: KOXAHOB@yandex.ru
НЕ НЕЙРОННАЯ СЕТЬ, А – НЕРВНАЯ ТКАНЬ!
Математические модели мозга, в виде нейронных сетей, зачастую игнорируют вклад глиоцитов, межклеточного матрикса и тканевой жидкости в работу мозга. Предпринята попытка дать биологическое обоснование надстройкам, корректирующим работу нейронной сети с помощью изменения величины синаптической задержки и скорости распространения возбуждения.
Ключевые слова: гликаны, глиоциты, синхронизация.
Введение
Несмотря на развитие автоматов, иногда заменяющих собой рабочего, что вдохновляет сторонников бионического моделирования мозга, успехи электрофизиологов, изучающих мозг, вдохновили McCalloch W.S., Pitts [1] на логическое исчисление идей, относящихся к нервной активности, а потом Фрэнка Розенблатта на создание персептрона, что и привело к конструированию биологически обоснованных искусственных нейронных сетей (БОИНС). Для обучения БОИНС используется метод коррекции ошибки, однако, с биологическим обоснованием функции УЧИТЕЛЬ имеются проблемы. Другая проблема в том, что непонятно: какие факторы значимы и включение их БОИНС обязательно, а какими можно пренебречь. Несмотря на критику нейронной доктрины[2], Wulfram Gerstner с соавторами [3] выпускает учебник по моделированию нейронных сетей, где развивающийся стохастический резонанс трактуется как основа познавательных способностей, если я правильно перевел cognition. H. Markram [4] делает более подробную модель, с включением астроцитов и данных коннектома, и симулирует на ней появление популяционных спайков, однако, в перспективе Blue Brain Project вырисовывается развитие стохастического резонанса в более сложной системе, а не качественного скачка в процессе познания работы мозга.
Экспериментаторы продолжают поставлять материал, который показывает, что мозг, т.е. прототип, гораздо сложней публикуемых моделей, которые описывают его работу. Jason J. Moore [5] показывает, что на дендритах формируются спайки, но если этот процесс можно описать, разбив реальный нейрон на совокупность формальных нейронов, то работа Кейли Балман[6], продемонстрировавшая субпопуляцию микроглии, ассоциированную с аксональным холмиком, поднимает вопрос о степени влияния этих клеток на активность нейронной сети в реальном мозге. Микроглия существует в резидентной и мобильной форме и способна к пролиферации, а, следовательно, к накоплению соматических мутаций. Смоделировать поведение микроглии можно, описав её как систему автономных модулей, а это уже другой принцип обработки информации о стимулах окружающей среды.
Как известно, стимулы окружающей среды, в зависимости от их физиологической значимости для организма, формируют образы в процессе консолидации памяти. Если кратковременная память зависит от электрофизиологической активности нейронов, чувствительна к электрошоковому воздействию и ингибиторам синтеза белка, то образы долговременной памяти могут храниться в течение всей жизни особи, и нечувствительны к вышеперечисленным перипетиям. Субстанция для хранения образов долговременной памяти неизвестна; физиологический механизм актуализации образов – неизвестен. Однако, известна система адаптивного иммунитета, способная к сохранению образа антигена на протяжении все жизни, и целесообразно выяснить степень гомологии между этими физиологическими системами.
Кибергены.
Разрабатывая формальную нейронную сеть, В.Филлипов столкнулся с необходимостью программного обращения к нейронам с целью корректировки синаптических весов и присвоил нейронам индексы, назвав их кибергенами. Похожая проблема была и А.Б. Когана: «Если нейронные ансамбли существуют, то они должны чем-то отличаться». Со времен Wilder Penfield известно, что нейроны как-то связаны с образами, а с помощью ф-МРТ иногда удается установить, какие образы актуализированы. Xu Liu с соавт.[7] удалось промаркировать нейроны, связанные с образом, а потом оптогенетическим воздействием актуализировать образ, это повтор достижений Пенфилда, или нечто большее? «То зевота нападёт, то охота пропадёт — что-нибудь непременно случается!», - это нормальная ситуация при тренировке абстрактного мышления, однако, потренировавшись, мы в состоянии рассортировать значимые и незначимые ассоциации связанные с образами.
Физиологический механизм консолидации памяти и актуализации образов должен иметь достаточную информационную емкость, и лабильность, сопоставимую с электрофизиологической активностью нейронов. Однако, ДНК, стабилизированная механизмами репарации, а так же непосредственно транскрибируемые и транслируемые с ней продукты, оказались заблокированы по математическим причинам, о которых я доложил на 5-ом съезде биофизиков России [8]. Кроме того, треть нейронов у мегафрагма мимерипинес [9] на стадии имаго (когда животное охотится, летает и размножается) теряют ядра. Получается, что ДНК нейронов в процессе консолидации памяти не всегда востребована. Более того, внутриклеточные субстанции отделены плазмолеммой от межклеточного сообщества, т.е. пространственно недосягаемы для контактного взаимодействия. Однако, кроме наследуемого информационного массива в виде ДНК, есть еще и информационный массив формируемой особью.
Гликаны состоят из шести различных мономеров, у которых есть шесть реакционноспособных связей. У электрического угря имеется 500 мономерных остатков в составе гликанов натриевого канала, но обычно мономерных остатков на натриевом канале в три раза меньше [10]. Подсчитаем количество вариантов структуры 6*6**166= 2,2193*10**258. Вроде бы много, но живыми организмами используется гораздо меньше структурных вариантов гликанов, т.е. структура находится под давлением отбора. Структура гликанов зависит протеолиза по анкерному домену фермента, в результате чего получаются растворимые изоферменты, у которых появляется возможность для взаимодействия с сайтами субстрата-акцептора, ранее недоступными для транслируемых мембранноассоциированных изоферментов. Какие факторы инициируют протеолиз неизвестно, возможно – функциональное состояние клетки. В частности, Mara Grube с соавторами [11] сообщила, что экзосомы, генерируемые тканью подвергшиеся гипоксии, меняют спектральные характеристики связанные с гликанами.
Отметим, что обработка нейронами, информации связанной с образом, относится к периоду кратковременной памяти, т.е. чувствительна к ингибиторам синтеза белка. Одновременно с синтезом белка происходит гликозилирование сайтов, связанных с контролем качества фолдинга, а непосредственно после этого происходят дальнейшие модификации гликанов, т.е. заблокировав синтез белка, мы так же заблокируем синтез гликанов, входящих в состав гликопротеинов.
Работа CAZy (Carbohydrate-Active Enzymes) [12] зависит от наличия субстрата-донора. В связи с этим, следует упомянуть о секреции в межклеточную жидкость гликозилтрансфераз, там субстраты-доноры не обнаружены в заметных количествах и отсутствуют ферменты для синтеза субстратов-доноров. В этом случае, клетки многоклеточных организмов явно отклоняются от эгоистической линии поведения, поскольку существует теоретическая возможность для направления CAZy в лизосомы, с целью пополнения внутриклеточного аминокислотного пула. Вероятно, субстраты-доноры для CAZy искали недостаточно тщательно, несмотря на то, что особенности гликозилирования межклеточного матрикса [13], [14] влияют на выбор направления дифференцировки стволовыми клетками.
Гликаны располагаются на наружной части плазматической мембраны и могут передавать межклеточному сообществу сообщения о внутриклеточных событиях, произошедших в аппарате Гольджи и эндоплазматическом ретикулюме.
Чем считывается индекс кибергена?
Во-первых, изменение степени ветвления гликанов, которое зафиксировано для некоторых случаев неопластической трансформации клеток [15] [16] [17], может изменить локальную концентрацию углеводов и вызвать гель-золь фазовый переход в гликокаликсе. Массоперенос в геле менее эффективен, чем в золе, что сказывается на вольтамперных характеристиках ионных каналов, в том числе.
Во-вторых, из-за легкости конформационных изменений гликаны способны к взаимодействию с веществами, к которым нет специфических протеиновых рецепторов.
В-третьих, существуют лектины, вещества специфически связывающиеся с карбогидратами.
Воспользовавшись базой данных, любезно выложенной в сеть сотрудниками Стендфордского университета, я заметил специфическое повышение транскрипции siglec (Sialic acid-binding immunoglobulin-type lectin) в микроглиоцитарной популяции нервной ткани[18]. филогенетическое дерево белков иммуноглобулинового суперсемейства[19], к которым относится singlec распадается на две ветви: она работает в нервной системе, другая - в системе адаптивного иммунитета. Однако, количество вариантов singlec существенно меньше вариантов электрофизиологической активности нейронов, т.е. наличествует несовпадение систем по информационной емкости.
Система адаптивного иммунитета внезапно появилась у позвоночных, а другие животные, и ланцетник – тоже, успешно выживают без неё. Поскольку нервная система более древняя, то в ней можно поискать зачатки системы адаптивного иммунитета, предполагая, что эти зачатки имеют функциональную нагрузку, а потому не элиминированы эволюционным процессом. Так, реаранжировка иммуноглобулиновых генов является сложным механизмом, требующим нескольких мутаций для своего формирования, а, следовательно, и времени для эволюции и прототипа, из которого может сформироваться механизм. Я проверил rag1 и rag2 гены, но они оказались супрессированы в нейронах, олигодендроцитах, в клетках предшественниках, астроцитах, микроглиоцитах.

Рис.1 Фрагмент результатов обработки базы данных An RNA-Seq transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. Цветная заливка, маркирующая разрыв интерфейса, заменена буквой русского шрифта.
Сравнив уровни транскрипции интерлейкинов и рецепторов к ним, я заметил, что иногда интерлейкин синтезируется, а рецепторы к нему отсутствуют, или же наоборот рецепторы к интерлейкину имеются у клеток нервной ткани, а ни одна из субпопуляций представленных в базе данных не синтезирует интерлейкина. Т.е. приёмники информации имеются, а передатчиков не имеется, т.е. интерфейс разорван. Вероятно, важное функциональное звено ЦНС выбросили в мусор, проводя первичный отбор субклеточных популяций, и это вполне могли быть лимфоциты, число которых варьирует в ЦНС, а повышение уровня считается маркером скрытого воспалительного процесса.
Но, какие клетки были прототипами лимфоцитов в эволюционном процессе? Понятно, что при недостаточном количестве вариабельных генов, лимфоцитарное звено системы адаптивного иммунитета не будет эффективным, поскольку биологическое разнообразие патогенов, на этом этапе эволюции, уже достаточно велико.
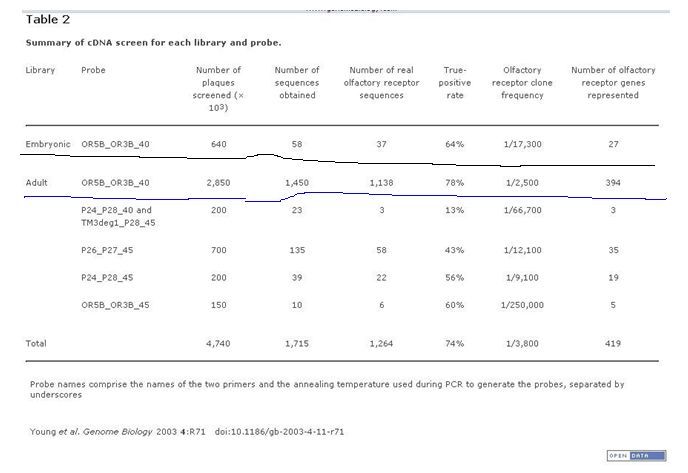
Табл.1 Цитировано [20]. Число фрагментов ДНК, кодирующих обонятельные рецепторы, у эмбриона и взрослой мыши существенно отличаются.
За неимением лучшего, перестройки в OR-семействе белков предлагаю считать функциональным (поскольку отсутствует гомология ДНК) прототипом реаранжировки иммуноглобулиновых генов, потому что:
- давление отбора направлено на расширение идиотипического многообразия транслируемых протеинов,
- сохранена пропорция одна клетка – один идиотип,
- количество уникальных последовательностей у взрослого животного больше, чем у эмбриона.
Если канцерогенез рассматривать как эволюционно незавершенный гистогенез, когда новообразование с полезной функцией снижает степень озлакачествленности в ряду поколений и становится новой тканью, то тогда формирование системы адаптивного иммунитета можно представить следующим образом:
-нейробластома, экспрессирующая обонятельные рецепторы определенного идиотипа, концентрирует на своей поверхности патоген
-фагоцитоз сконцентрированного патогена оказался более эффективным
-рецепторы лимфоцитов эволюционируют из кадхеринов, которые сначала выполняли вспомогательную роль в удержании патогена, а в процессе эволюции, уже в составе системы адаптивного иммунитета, постепенно увеличили свою вариабельность, аффинитет и селективность экпрессии кодирующих их генов. Что согласуется с гипотезой Галлактионова В.Г. [21]
А теперь обратим внимание на миелиновые оболочки[22]. У зверей они представлены концентрической спиралью, а у первичноротых расположены вдоль нейрита, по факту – наличествует конвергенция, зачем? Учтем, что синхронизация постсинаптических потенциалов на дендритах интегративного нейрона может способствовать формированию потенциала действия, как осуществить синхронизацию технологически?
Если менять скорость передачи ПД в рецептивном поле нейрона, то синхронизации добиться можно, а именно, адгезией глиальной клетки на нейрите, что вызовет сальтоторную передачу возбуждения на этом участке нейрита. Блокирование каналов за счет образования супрамолекулярных комплексов между карбоновыми группировками ацетилнейраминовой кислоты и аминогруппами полиаминов так же может вызвать сальтоторную передачу возбуждения на молекулярном уровне. То есть, что-то должно незначительно менять скорость проведения потенциала действия, прежде чем эволюционным процессом будет поддержана тенденция, которая и привела к формированию миелиновых оболочек у неродственных организмов. Незначительная разница по времени может быть существенно увеличена за счет циклов реверберации, если циклы реверберации возбуждения всё-таки имеются.
Заключение.
На основании вышеизложенного, можно сделать следующие выводы:
1. В гликанах могут происходить структурные изменения, связанные с электрофизиологической активностью нейрона.
2. Микроглиоциты могут использовать siglec при мониторинге функционального состояния нейрона. Возникает вопрос об использовании этой функциональной связи в процессе синаптического прунинга.
3. Концепция нейронной сети недостаточна для моделирования работы мозга, поскольку не учитывает: а) нейроглиальные взаимодействия, функционирующие как система автономных модулей, б) процессы спилловера, работающие как локальные аналоговые вычислители в среде межклеточного матрикса.
4. Предложена эвристическая модель работы нервной ткани, с учетом экспериментально подтвержденной вариации времени синаптической задержки в пределах 10% [23].
Допустим, действие, вызываемое мотонейроном, не приводит к желаемому результату, из-за этого меняется состав коктейля нейромедиаторов в межклеточной среде, что изменяет время синаптической задержки в цикле реверберации №3. В результате десинхронизации в циклах реверберации возбуждения №3 и №2, исполнительный мотонейрон не получает синхронного поступления возбуждающих постсинаптических потенциалов от ассоциативного нейрона и нейрона цикла №2 (назовем его гностическим). Какая нервная цепочка возбудится в результате синхронной работы мотивирующего (цикл реверберации№3) и поискового (цикл реверберации №1) нейронов – про то не известно, но есть вероятность, что использование оператора CASE (уже биологически обусловленного) может снизить активность в мотивирующих нейронах. Кстати, расчетная частота возбуждения нейрона в гипотетических циклах реверберации возбуждения случайно совпала с гамма-ритмом, который физиологи связывают с напряженной умственной деятельностью.
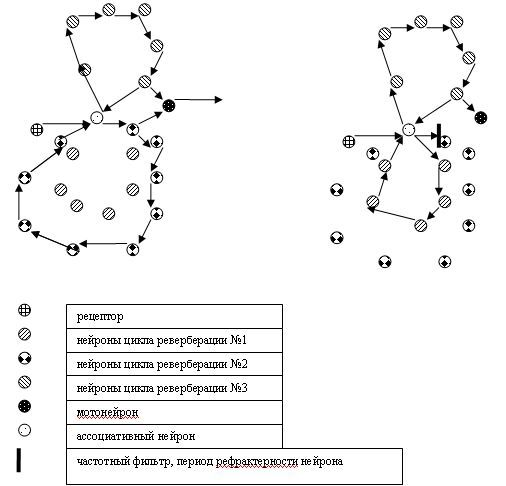
Рис.2 Изменение структуры нейронных ансамблей при вариации времени синаптической задержки. Нейроны и нейриты, не имеющие отношения к циклам реверберации возбуждения, проигнорированы.
Список литературы
1 McCalloch W.S., Pitts W. «A logical calculus of the ideas immanent in nervous activity» // Bull. Math. Biophys. – 1943. – v.5. – pp.115–133.
2 Bullock T.M et al «The Neuron Doctrine, Redux»// Science 310 4 nov. 2005 791(3)
3 Gerstner Wulfram et al. «Neuronal dynamics : from single neurons to networks and models of cognition» - Cambridge University Press 2014г. 590стр.
4 Markram H. et al. «Reconstruction and Simulation of Neocortical /Microcircuitry» //Cell Volume 163, Issue 2, p456–492, 8 October 2015
5 Moore J.J. et al «Dynamics of Cortical Dendritic Membrane Potential and Spikes in Freely Behaving Rats» // bioRxiv preprint first posted online Dec. 28, 2016; doi: http://dx.doi.org/10.1101/096941.
6 Baalman K. et al. «Axon Initial Segment–Associated Microglia» //The Journal of Neuroscience, February 4, 2015 • 35(5):2283–2292 • 2283
7 Xu Liu et al «Inception of a false memory by optogenetic manipulation of a hippocampal memory engram»// Philos Trans R Soc Lond B Biol Sci. 2014 Jan 5; 369(1633):
8 Коханов В.С. «Поведение, сознание. Бритва Оккама. Бактерия, антибиотик, хищник»// 5-ый съезд биофизиков России материалы докладов т.2 стр. 318
9 Polilov A.A. «The smallest insects evolve anucleate neurons»// Arthropod Struct Dev. Volume 41, Issue 1, January 2012, Pages 29–34
10 Duclohier H. Structure–function studies on the voltage-gated sodium channel // Biochimica et Biophysica Acta (BBA) – Biomembranes Volume 1788, Issue 11, November 2009, Pages 2374–2379
11 Grube M., Shvirksts K., Abols A., Line A. (2015) Evaluation of cancer-derived exosomes by FT-IR spectroscopy,// 10th Workshop FT-IR Spectroscopy in Microbiological and Medical Diagnostics, October 15-16, Berlin, Germany, Proceedings, p.89,
12 Carbohydrate-Active Enzymes http://www.cazy.org/
13 Nobuaki Maeda «Functional Roles of Chondroitin Sulfate in Neural Network Formation» (Tokyo Metropolitan Institute for Neuroscience, Department of Developmental Neuroscience) http://www.glycoforu…-A01E.html
14 Tung-Ying Lu et al «Repopulation of decellularized mouse heart with human induced pluripotent stem cell-derived cardiovascular progenitor cells» // Nature Communications 4, Published online: 13 August 2013 Article number: 2307
15 Ajit Varki et al. «Essentials of Glycobiology» 2nd edition - Cold Spring Harbor Laboratory Press; 2009. 784 pages ISBN-13: 9780879697709
16 Zhengyi Ye and Jamey D. Marth «N-glycan branching requirement in neuronal and postnatal viability» // Glycobiology. 2004 Jun;14(6):547-58. Epub 2004 Mar 24.
17 Scott H. and Panin V. «The role of protein N-glycosylation in neural transmission» // Glycobiology vol. 24 no. 5 pp. 407–417, 2014
18 An RNA-Seq transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex http://web.stanford.…naseq.html
19 Kyoto Encyclopedia of Genes and Genomes http://www.kegg.jp/k…uperfamily
20 Young J.M. et al «Odorant receptor expressed sequence tags demonstrate olfactory expression of over 400 genes, extensive alternate splicing and unequal expression levels» // Genome Biol. 2003;4(11):R71 http:/genomebiology.biomedcentral.com/articles/10.1186/gb-2003-4-11-r71
21 Галактионов В.Г. «Эволюционная иммунология» – М.: Академкнига, 2005. – 408 с
22 Kettenmann H. and Ransom B. «Neuroglia» Third Edition - Oxford University Press 2012 год 995 стр.
23 Jen-Wei Lin and Faber D. «Modulation of synaptic delay during synaptic plasticity» // Trends in Neurosciences Vol.25 No.9 September 2002 p 449
P.S.
Рад, что теоретическое обоснование получило дальнейшее развитие при разработке программного обеспечения, представленного сотрудниками института Прикладной физики Яхно В.Г. с соавторами в докладе на конференции «Нейроинформатика 2017»

Отредактировано: VladimirKox - 06 янв 2018 в 10:23
- +0.00 / 0
-
Малая Вишера
Карма: -21.94
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Регистрация: 12.01.2017
Сообщений: 1,826
Читатели: 1
Ну, вы, блин даете.
И все же нет никакого смысла в пошаговом повторении всех механизмов и процессов в человеческом мозге. Это ничего не даст с точки зрения создания искуственного разума.
Культ карго в чистом виде.
Как пример - миллиарды лет эволюции живых существ так и не привели к появлению колеса. Природа не смогла, хотя информация вот она, бери. А человек смог.
На пальцах модель сможете описать?
Знания они не в институте и монография, а в голове. Никакие костыльки человеку не нужны.
Для познания вселенной и природы разума у человека все есть от рождения.
Человек способен познать и описать вселенную голый, на голой земле.
И все же нет никакого смысла в пошаговом повторении всех механизмов и процессов в человеческом мозге. Это ничего не даст с точки зрения создания искуственного разума.
Культ карго в чистом виде.
Как пример - миллиарды лет эволюции живых существ так и не привели к появлению колеса. Природа не смогла, хотя информация вот она, бери. А человек смог.
На пальцах модель сможете описать?
Знания они не в институте и монография, а в голове. Никакие костыльки человеку не нужны.
Для познания вселенной и природы разума у человека все есть от рождения.
Человек способен познать и описать вселенную голый, на голой земле.
"Мне плевать на вас ублюдки.
Я анархо-аморал." (С)
Я анархо-аморал." (С)
- 0.00 / 2
-
Карма: +4.92
Регистрация: 01.04.2011
Сообщений: 1,430
Читатели: 0
Регистрация: 01.04.2011
Сообщений: 1,430
Читатели: 0
Цитата: mark.76 от 06.01.2018 12:18:29И все же нет никакого смысла в пошаговом повторении всех механизмов и процессов в человеческом мозге. Это ничего не даст с точки зрения создания искуственного разума.
Культ карго в чистом виде.
У химиков органиков есть традиция, после того как они устанавливают структурную формулу вещества, они это вещество пытаются получить искуственно, а потом сравнивают свойства природного вещества и синтетического аналога. Так они проверяют, что уже узнали, а что ещё не узнали.
Расписать онтологию мозга будет полезно для физиологов, медицины, фармакологов. Для инженера онтология - справочная информация, что захочет - то и использует в своих поделках. Нам не нужно, чтоб самолет махал крыльями, достаточно того чтоб он летал. Но я не уверен, что сейчас инженеры используют профиль крыла птицы, а не что-то другое. У Генри Маркрама, с финансированием 1 миллиард евро, синхронная активность в нейронной сети гаснет через 160 милисекунд( и хоронит гипотезу Савельева память=эл.физ. активность НС ), у речного угря месторасположение Саргасова моря хранится всю жизнь и активизируется в нужный момент.
Вам не интересно из чего состоит учитель=корректор нейронной сети? Вам не интересно как особь ставит перед собой исследовательские задачи?
P.S.
Герман Греф назвал голосового помощника Алису глупой: "С ней невозможно поговорить". Алиса ответила:"Рыбак рыбака..." Что ж, пожелаем ему достаточного видеокарт, для изготовления среды обучения для нейронной сети.
P.P.S.
Лженаука отличается тем, что игнорирует накопленный массив знаний.
Отредактировано: VladimirKox - 06 янв 2018 в 20:06
- +0.06 / 2
-
Сейчас на ветке:
2,
Модераторов: 0,
Пользователей: 0,
Гостей: 1,
Ботов: 1