Вселенная, жизнь, разум...
105,189
250

41 год
Карма: +0.14
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Цитата: planus от 03.10.2017 22:11:20Ресурсами занимались товарищи из нашей солнечной системы. А не с других систем или галактик. Это имеет подтверждение в легендах и раскопках. А вот всякие "серые" и прочие - скорее всего не прилетали, а воздействовали ментально на мозг издалека.
-
Это да. Золото довольно редкий продукт взрыва нейтронных звёзд или " ...происхождение элементов тяжёлых, таких как золото, платина и уран, до сих пор для учёных во многом остаётся загадкой".Думаю что Земля как-то ухитрилась попасть под метеоритную бомбардировку золотыми метеоритами. И содержание золота в коре Земли во много тысяч раз превышает расчётные варианты.

Про это и Сетчин пишет, что золотодобытчики с Нибиру а она в солнечной системе, но в мифах описываются разные расы и я сомневаюсь что далекая от Солнца планета способна поддерживать жизнь даже с золотом. Невозможно представить что древние цивилизации не смогли освоить межзвездные перелеты, так что за золотом на землю летают наши соседи(добрые и недобрые) с других звезд.
Золота на Земле очень много в тысячи раз больше - не думаю, просто много, достаточно много, и удобно добывать - люди сами добывают - на Венере попробуй добыть пусть там золотые горы или даже на Марсе
воздуха нет, воды нет, населения нет...
41 год
Карма: +0.14
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Цитата: Свой от 03.10.2017 22:25:341) В данном вопросе интересны только желтые карлики. Мы ведь говорим про планетарные цивилизации?
2) Цивилизаций, отставших от нас и наших ровесников на 500 тыс лет не возникнет. Мы будем осваивать их планеты еще в эпоху динозавров.
Цивилизаций второй волны не возникнет. Кто не успел – тот опоздал.
1. А красные карлики, фильм Хищник смотрели? Хищник с планеты с большой гравитацией, плотной теплой атмосферой но тусклым красным освещением.
Из семи известных на сегодня планет системы три находятся в обитаемой зоне TRAPPIST-1: e, f и g звезда красный карлик. Его планета примерно такая - освещение как на Марсе в 2-3 раза слабее, гравитация примерно в 3 раза сильнее, давление 2-3 атмосферы, она относительно далеко от звезды но плотная атмосфера создает парниковый эффект поэтому средняя температура примерно +30( у земли +15), планета Хищника покрыта жаркими красными джунглями полными жизни - это все у Красного Карлика.
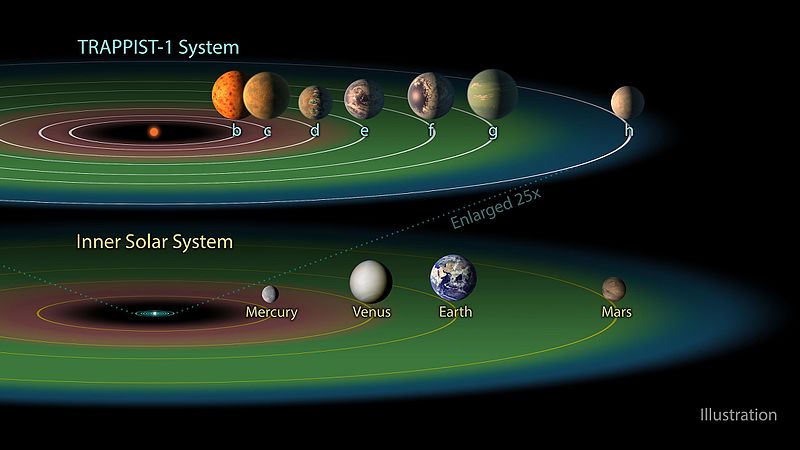
2. Не понимаю почему, в системе Альфа Центавра разумная жизнь могла возникнуть на 500 млн лет раньше, а ведь есть желтые карлики на миллиарды лет старше Солнца.
3. Опоздали наверное мы.
Отредактировано: Слава333 - 03 окт 2017 23:30:03
41 год
Карма: +0.14
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Цитата: planus от 03.10.2017 22:53:57А без звёздного тепла и света как они вообще долетели (и за какое время) до солнечной системы ?
-
Там жить возможно было лишь под планетной корой. Куда не дойдёт космический холод и радиация.
-
Долетели, есть технологии межзвездных перелетов, керосин(ракетное топливо) для этого жечь не нужно, по описаниям есть антигравитационные двигатели - летающие города и пр.
Да жить в глубоких пещерах можно и на Плутоне, но возникает пару вопросов.
1.Как могла появиться жизнь, тем более разумная жизнь, на такой страшно холодной планете?
2. Зачем там вообще жить если в системе есть Земля и есть возможность туда переселиться?
41 год
Карма: +0.14
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Цитата: planus от 03.10.2017 23:46:251. Она не была страшно холодной. Она тоже грелась своей звездочкой.Это потом она ушла с орбиты и полетела к нам в гости. Беженцы короче..
-
2. Земля очень проблемная для метеоритных ударов. Наверное просчитали эти орбиты и время попадания. Плюс на Земле ОКЕАН. Частые потопы. Помните как Шумеры писали: "Шумерская табличка о потопе считается документом и имеет огромную ценность для истории человечества. В ней не хватает верхней части таблички, которая содержала 37 строк из древнешумерского эпоса о потопе. Именно в этой части, видимо, говорилось о причинах, по которым боги решили погубить людей. Видимый текст начинается с желания верховного бога спасти человечество от полного вымирания. Им движет вера в то, что к людям вернется почитание тех, кто их создал."

1. Тогда беженцы могли расположиться на хорошей орбите поближе к Солнцу. И как они вообще стартовали от своей звезды и целой планетой долетели к нам, не понимаю.
2. Метеориты проблема для всех планет в солнечной системе но не такая уж огромная проблема, последний по настоящему большой метеорит упал 75 миллионов лет назад, неужели лучше жить при -250 без атмосферы чем в тепле на Земле боясь метеоритов?
3. Потопы - сильно преувеличены, к тому же есть горы, живи на 100-1000 м выше уровня моря и радуйся
Отредактировано: Слава333 - 03 окт 2017 23:56:14

Potsdam
57 лет
Карма: +153.88
Регистрация: 28.09.2012
Сообщений: 5,615
Читатели: 4
Регистрация: 28.09.2012
Сообщений: 5,615
Читатели: 4
Цитата: Слава333 от 03.10.2017 21:26:20Отчасти согласен, кислорода было больше и плотность атмосферы была раза в 2 выше, но все это явно недостаточно чтобы поднять в воздух такое чудовище
В Румынии палеонтологи обнаружили останки гигантского птеродактиля, размах крыльев которого достигал 16 метров а вес до 300 кг.
Самая большая летающая птица сегодня Кондор весит до 15 кг - в 20 раз меньше!

ну давайте посмотрим, что мы имеем в авиастроении.
мотопланер 300 кг - двигатель 16 кВт на взлете - 60 метров разбег, 3 кВт в наборе и 2 кВт в горизонтальном полете, скорость до 240 км/час. дельтаплан - взлет с возвышенности и использование термиков, безмоторный полет.
комбинируем и получаем 300 килограммовый птеродактиль взлетает с возвышенности и развивая 3 кВт вполне сносно может набирать высоту даже в нашей атмосфере. для сравнения - тренированный велосипедист (пусть будет 75 кг) развивает долговременную мощность 500-700 Ватт и 1,5-2 кВт пиковой.
вывод - птеродактиль мог-бы летать в нынешней атмосфере. разумеется с ровной полощадки он бы не взлетел. разве что мог бы развивать киловатт 12 пиковой мощности. можно и этого добиться. эволюция однако.
No pasarán ... fama a los héroes ... bommbass para siempre ...
учиться военному делу настоящим образом ... и скажите уже государю, что у англичан ружья кирпичом не чистят ... пусть чтобы и у нас не чистили, а то они стрелять не годятся ...
учиться военному делу настоящим образом ... и скажите уже государю, что у англичан ружья кирпичом не чистят ... пусть чтобы и у нас не чистили, а то они стрелять не годятся ...
41 год
Карма: +0.14
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Цитата: planus от 04.10.2017 00:21:511. Управлять такими массами как их планета беженцы скорее всего не умеют.А то бы не стали перпендикулярной планетой.
2. А Мамонты сами себя недавно "в вечную мерзлоту (шутники эти "учёные")зарыли ?
3. Озеро Титикака.
1. Вы писали что беженцы прилетели с целой своей планетой, вот я и подумал что это возможно только если они могли управлять её движением.
Не могу представить естественный процесс в ходе которого населенная планета сойдет с орбиты возле чужой звезды и полетит к другой, в данном случае к Солнцу, к тому же дорога неизбежно займет сотни тысяч лет в холоде и мраке межзвездного пространства.
2. Перемены климата - ледниковый период, не обязательно причиной гибели мамонтов была космическая катастрофа, слоны-бегемоты ведь выжили.
3. А что с этим озером - до него доставал "потоп"? Как то сомнительно, тогда бы все крупные формы жизни на Земле "слоны-бегемоты" погибли.
41 год
Карма: +0.14
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Регистрация: 01.09.2012
Сообщений: 370
Читатели: 0
Цитата: planus от 05.10.2017 19:14:06В отличии от Ситчина и прочих, я подозреваю, что это наша планета.
-
С озером всё просто. Там солёная океанская вода и океанская фауна. Высота над уровнем моря почти 4 км. Это самое высокогорное озеро.
Есть 2 варианта:
1. Потоп принёс воду и обитателей в пустую от воды котловину. Волна выше 4 км.
2. Это было побережье океана. А потом оно вдруг без катапульты поднялось на 4 км.
1. Эта гипотеза выглядит правдоподобнее - планета погасшей звезды-спутника Солнца.
2. Титикака пресноводное озеро хотя с повышенной соленостью, флора и фауна там вроде пресноводная.
Я все время повторяю - никаких планетарных катастроф(потопов, метеоритов, вулканических зим) за последние миллионы лет не было иначе все крупные животные неминуемо бы погибли, последняя была 75 млн лет назад когда вымерли динозавры - я имел в виду на Землю тогда в последний раз упал большой метеорит, с тех пор падали относительно мелкие. А что на Юпитер падают кометы - что тут удивительного, планета очень большая и находится возле пояса астероидов.
Отредактировано: Слава333 - 05 окт 2017 19:40:46


Москва
Карма: +193.83
Регистрация: 21.03.2013
Сообщений: 27,810
Читатели: 7
Регистрация: 21.03.2013
Сообщений: 27,810
Читатели: 7
Цитата: planus от 05.10.2017 19:14:06В отличии от Ситчина и прочих, я подозреваю, что это наша планета.
-
С озером всё просто. Там солёная океанская вода и океанская фауна. Высота над уровнем моря почти 4 км. Это самое высокогорное озеро.
Есть 2 варианта:
1. Потоп принёс воду и обитателей в пустую от воды котловину. Волна выше 4 км.
2. Это было побережье океана. А потом оно вдруг без катапульты поднялось на 4 км.
-
Вы какой бы вариант выбрали ?

Метеорит 12 тыс лет назад проломил земную кору и обломки одними краями поднялись, а другими опустились. Заодно вызвал Великий Потоп в виде гигантского цунами в других частях света. Испарил гигантское количество воды, после чего она лилась с неба ещё месяц )
Так была уничтожена Предыдущая Цивилизация.
Отредактировано: slavae - 06 окт 2017 07:38:12
Империя - это мир, и этой идеологии достаточно. Мы живём в самой лучшей стране в мире и все нам завидуют.
Одушевлённое Одевают, Неодушевлённое Надевают.
Одушевлённое Одевают, Неодушевлённое Надевают.
Москва
Карма: +193.83
Регистрация: 21.03.2013
Сообщений: 27,810
Читатели: 7
Регистрация: 21.03.2013
Сообщений: 27,810
Читатели: 7
Цитата: planus от 06.10.2017 15:49:59Но бабахнул он сильно далеко от этого озера чтобы поднять его на 4 км..
В феврале 2012 года Национальная академия наук США опубликовала доклад, подтверждающий падение метеорита в Мексике 13 тыс. лет назад
Империя - это мир, и этой идеологии достаточно. Мы живём в самой лучшей стране в мире и все нам завидуют.
Одушевлённое Одевают, Неодушевлённое Надевают.
Одушевлённое Одевают, Неодушевлённое Надевают.
56 лет
Карма: +7.60
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Цитата: ДядяВася от 23.06.2017 20:27:55Конспирология это конечно интересно, но:
1. Более менее за мощную нейтронную вспышку можно принять только Чернобыль (по сравнению с нормально работающими реакторами), остальное семечки.
Не было никакой нейтронной вспышки в чернобыле -была пароциркониевая реакция...Я это вам как обитатель киева 1986-1987 года говорю - писька у меня нормально стоит, как и у многих других обитателей
Цитата
2. Мощность атомных - термоядерных взрывов (в т.ч. и по нейтрино) на много порядков превосходят реакторы. А их в ходе мирных и немирных взрывов порядка 1000 с каждой стороны (США - СССР), ну и других, по мелочам.
3. Мощность солнечных нейтрино в "сиксилион - сиксилионов" превышает земные.
Выводы делайте сами.
56 лет
Карма: +7.60
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Цитата: Слава333 от 02.10.2017 21:34:13Предлагаю интересную тему - на другой ветке некоторые засомневались что в Галактике есть хоть одна Цивилизация способная летать к другим звездам, как вы думаете есть ли такие и вообще сколько Цивилизаций в нашей Галактике?
Вопрос - Предоставьте, пожалуйста, пруф на существование цивилизации, летающей на десятки и более световых лет?
Мой ответ - На Землю много прилетало, разные расы - об этом говорят мифы многих народов, и наверно до сих пор летают.
Или вы думаете что из многих миллионов цивилизаций нашей Галактики ни одна не освоила межзвездные перелеты, все сидят по своим планетам и никуда не летают?

Надо славу в бан отправлять ...вечный... шо не мучилсо...и флейма не созавал, за одно и артефакты пока в бане подыскал
56 лет
Карма: +7.60
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Регистрация: 16.07.2012
Сообщений: 3,879
Читатели: 1
Аккаунт заблокирован
Цитата: Свой от 03.10.2017 12:14:36.
А чего тут обсуждать?
Это голимая арифметика.
Если верить аббату Жоржу Леметру и папской Академии Наук, Бог создал Вселенную 14.5 млрд лет назад. Но на тот момент это был чисто водородный мир.
все смешалст в доме облонских... леметр действительно автор гипотезы большого взрыва, но из данных хаббла лет этих было не 14,4 а около 2, что вызвало настоящий батхерт, так как не более чем через 10 лер ужо имелись оценки возраста зесли по радиоизотопному методу в боле чем 4 миллиарда лет
Цитата
+1 млрд – сформировались первые звезды
+9 млрд – взорвались первые звезды, и в процессе этого чуда прошли сложные реакции ядерного синтеза.
шо то мне кается 8 миллиардов лет на выгорание первых ,(а поэтому мало металлизированных звезд черезвычайно долго, учитывая чир они должно быть очень массивные[/quote]
+1 млрд – сформировались звезды второй волны и первые планеты с таблицей Менделеева
Возраст Земли – 4.5 млрд лет. Это значит, что она гарантированно находится в первой волне цивилизаций, имеющих доступ к полному спектру таблицы Менделеева.
Эволюция, скорее всего, везде идет примерно с равной скоростью.
.
Диагноз:
Вероятность того, что где-то во вселенной существуют похожие на нас цивилизации, ЗНАЧИТЕЛЬНО обогнавшие нас в развитии – пренебрежительно мала.
Возможно существование "водородных цивилизаций". Но мы вряд ли способны отличить их от природных явлений.[/quote]
не следует никак - ибо уникальность химии, условий формироваия, и размеры вселенной не оценимы...Все это вербальные оценки верующих...

Москва
20 лет
Карма: +282.16
Регистрация: 22.07.2010
Сообщений: 5,891
Читатели: 7
Регистрация: 22.07.2010
Сообщений: 5,891
Читатели: 7
Цитата: stranger1234 от 26.10.2017 00:31:57Не было никакой нейтронной вспышки в чернобыле -была пароциркониевая реакция...
Вы путаете причину взрыва реактора (вследствие пароциркониевой реакции) с причиной вызвавшей пароциркониевую реакцию.
Причиной парациркониевой реакции являлся неконтролируемый скачёк (на пару порядков) мощности реактора (нейтронная вспышка) вследствие которой произошло плавление топлива - пароциркониевая реакция - водород - взрыв водорода.

Южный Урал
Карма: +54.25
Регистрация: 14.02.2017
Сообщений: 1,070
Читатели: 3
Регистрация: 14.02.2017
Сообщений: 1,070
Читатели: 3
Долгосрочный эволюционный эксперимент на бактериях Escherichia coli, продолжающийся уже почти 30 лет, дал новые неожиданные результаты. Секвенирование ДНК бактерий из замороженной «ископаемой летописи» эксперимента показало, что за 60 000 поколений эволюция подопытных популяций не прекратилась и даже не замедлилась. Как минимум в 9 популяциях из 12 произошла экологическая диверсификация: исходная монокультура подразделилась на клады, связанные экологическими взаимодействиями и не вытесняющие друг друга. Внутри каждой клады эволюция продолжается полным ходом, причем дальнейшие изменения направляются как предшествующей эволюционной историей, так и меняющейся экологической обстановкой. Таким образом, эволюция «перехитрила» исследователей, надеявшихся изучить действие мутаций и отбора в ходе адаптации к стабильным условиям в предельно простой искусственной системе.
Скрытый текст
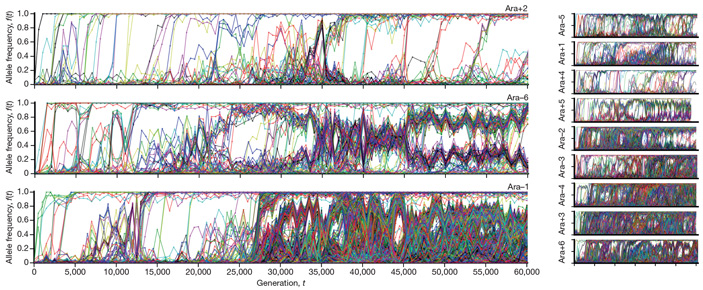

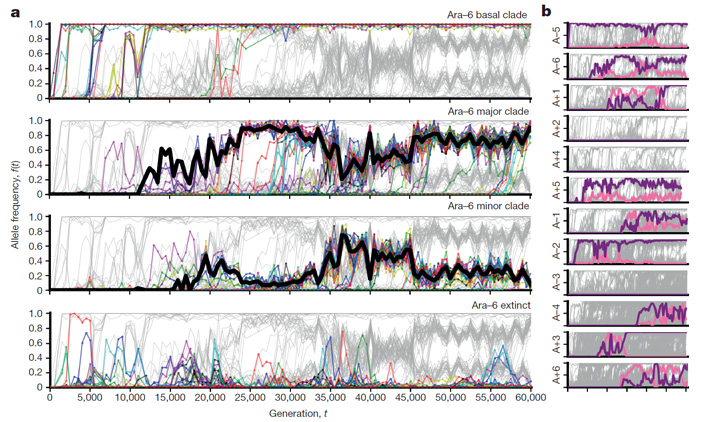
Статистический анализ распределения мутаций во времени показал, что в одних генах мутации преимущественно фиксировались в начале эксперимента (на ранних этапах адаптации), тогда как в других генах мутации начали фиксироваться лишь на поздних этапах. Это объясняется тремя причинами, причем все три, по мнению авторов, реально работают в ходе эксперимента (строго доказать это пока нельзя, но есть косвенные статистические аргументы в пользу реальности всех трех причин).
Во-первых, мутации в некоторых генах наиболее выгодны (дают наибольшую прибавку приспособленности) для исходного генотипа, и поэтому такие мутации фиксируются первыми, вытесняя в ходе клональной интерференции другие, менее полезные мутации в других генах. Эти последние начинают распространяться позже, когда первая порция самых «очевидных» полезных мутаций уже зафиксировалась.
Во-вторых, мутации, зафиксировавшиеся ранее, влияют на полезность или вредность мутаций, появляющихся позднее (влияние одних генетических вариантов на фенотипические проявления, в том числе на полезность, других называют эпистазом, см.: Epistasis). Поэтому некоторые мутации становятся полезными и получают шанс зафиксироваться только после того, как благодаря другим мутациям сформируется подходящий генетический контекст (см.: Эволюция белков сдерживается низкой проходимостью ландшафта приспособленности, «Элементы», 09.02.2015). Именно так обстояло дело с мутацией, благодаря которой бактерии из популяции Ara-3 получили возможность питаться цитратом (см.: В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшества, «Элементы», 25.09.2012).
В-третьих, формирующиеся экологические взаимодействия между кладами в корне меняют «правила игры», заставляя бактерий приспосабливаться уже не к стабильной и предельно простой среде (как было задумано исследователями), а к динамичному биотическому окружению. Это значит, что направленность отбора всё время меняется, и поэтому мутации, полезные для данной клады в данный момент, вовсе не обязательно будут полезными в другое время или для других клад.
Таким образом, долгосрочный эволюционный эксперимент опроверг чрезмерно упрощенные представления о том, как должна идти адаптация бесполой популяции к стабильным условиям среды. Ничего похожего на замедление и остановку адаптивной эволюции по мере приближения к оптимуму (пику на ландшафте приспособленности) не наблюдается, запас потенциально полезных мутаций не исчерпывается, и даже темп их накопления практически не снижается. Вместо этого мы видим самопроизвольное усложнение эволюционирующего сообщества, которое из монокультуры превращается в экосистему с подразделенными нишами и коэволюционирующими кладами и явно не собирается в обозримом будущем переходить в состояние эволюционного стазиса. Так что Лесли Оргел был, конечно, прав насчет того, кто умнее — эволюция или теоретики, считающие, что всё про нее знают.
Источник: Benjamin H. Good, Michael J. McDonald, Jeffrey E. Barrick, Richard E. Lenski & Michael M. Desai. The dynamics of molecular evolution over 60,000 generations // Nature. Published online 18 October 2017. DOI: 10.1038/nature24287.
Александр Марков
Будет али нет, а ты паши да сей. В лучшее верить надо!


Moskow
Карма: +1.30
Регистрация: 03.02.2010
Сообщений: 2,478
Читатели: 0
Регистрация: 03.02.2010
Сообщений: 2,478
Читатели: 0
Физикам из ЦЕРНа удалось проверить со сверхвысокой точностью закон CPT-симметрии для антиматерии и материи. Оказалось что они одинаковые с точностью, большей, чем две части на десять в минус десятой степени.
Т.е. для тех, кто понимает о чём я пишу - полное доминирование материи над антиматерией нарушением симметрии объяснить больше не получается, в момент Большого Взрыва все частицы должны были провзаимодействовать между собой и на выходе после аннигиляции остались бы одни фотоны и нейтрино... Нашего мира попросту бы не существовало. Тупик.
https://zen.yandex.r…2c3ddf8f07
Т.е. для тех, кто понимает о чём я пишу - полное доминирование материи над антиматерией нарушением симметрии объяснить больше не получается, в момент Большого Взрыва все частицы должны были провзаимодействовать между собой и на выходе после аннигиляции остались бы одни фотоны и нейтрино... Нашего мира попросту бы не существовало. Тупик.
https://zen.yandex.r…2c3ddf8f07
Большая буква Z

55 лет
Карма: +331.47
Регистрация: 20.11.2008
Сообщений: 27,590
Читатели: 53
Глобальный Модератор
Регистрация: 20.11.2008
Сообщений: 27,590
Читатели: 53
Глобальный Модератор
Цитата: Гарик от 06.04.2018 11:38:26Т.е. для тех, кто понимает о чём я пишу - полное доминирование материи над антиматерией нарушением симметрии объяснить больше не получается, в момент Большого Взрыва все частицы должны были провзаимодействовать между собой и на выходе после аннигиляции остались бы одни фотоны и нейтрино... Нашего мира попросту бы не существовало. Тупик.
А уже достаточно давно и не пытаются объяснять нарушением СРТ симметрии. Теперь применяют другое объяснение. Поле, из энергии которого рождались частицы, возникло (выкристализовалось, выделилось из симметрии/взаимодействия более высокого порядка) с неким случайным отклонением от минимума энергии. Возникло бы в минимуме, никаких частиц не было бы вообще. После чего совершило ряд постепенно затухающих колебаний, как в LC контуре при этом роль активного сопротивления, рассеивающего энергию и в конце концов прекращающего колебания, играло образование этих самых частиц. Но если обмотка в катушке индуктивности нагревается одинаково, независимо от направления тока, то здесь, когда "ток двигался слева направо" рождались частицы, а когда "справа налево" - античастицы. И в каждом цикле энергетика образования частиц была чуть выше, чем для античастиц. Точнее ту ветку, на которой случайно оказался первоначальный потенциал поля, мы сегодня называем частицами.
Ну или как в механическом маятнике. Если изначально мы его отклонили влево, то из за трения в подвесе каждая правая ветка колебаний будет чуть меньше, чем предшествующая ей левая. И сумма левых отклонений в конце немного превзойдёт сумму правых.
Отредактировано: Senya - 06 апр 2018 14:01:38
"Иван Грозный помещает на рабочий стол полученный от хана ярлык."(с) Не моё.
Карма: +53.31
Регистрация: 24.07.2008
Сообщений: 4,224
Читатели: 0
Регистрация: 24.07.2008
Сообщений: 4,224
Читатели: 0
Ну а что, подобное возможно. Слишком велика пропасть между звездами. Без "проколов пространства" пока разумных методов межзвездных перелетов нет.
Еще вариант, кстати, объясняющий знаменитый парадокс Ферми: на определенном уровне развития цивилизация неизбежно трансформируется в нечто, совершенно отличное от привычной нам формы - с материальными телами, традиционной техникой и технологией.
Еще вариант, кстати, объясняющий знаменитый парадокс Ферми: на определенном уровне развития цивилизация неизбежно трансформируется в нечто, совершенно отличное от привычной нам формы - с материальными телами, традиционной техникой и технологией.

Красногорск
73 года
Карма: +0.17
Регистрация: 27.09.2015
Сообщений: 7,486
Читатели: 6
Регистрация: 27.09.2015
Сообщений: 7,486
Читатели: 6
Цитата: Longspig от 08.04.2018 15:28:59Ежели кто "прошел мимо" - аккурат в тему (в качестве воскресного оффтопа)
Кэмпбелл Джон. Трансплутон
Так в этом видимо и есть Квинтэссенция
Красногорск
73 года
Карма: +0.17
Регистрация: 27.09.2015
Сообщений: 7,486
Читатели: 6
Регистрация: 27.09.2015
Сообщений: 7,486
Читатели: 6

Пока факт существования жизни за пределами Земли не установлен. Если говорить о солнечной системе, то в ней, по моему мнению, нет жизни. Единственный маловероятный сценарий – существование жизни на Марсе. Ученые предполагают, что глубоко в грунте Марса могли остаться какие-то организмы, если ранее на Марсе вообще была жизнь.
Дело в том, что примерно миллиард лет назад Марс обладал атмосферой и водой, и там могла зародиться жизнь. Ученые считают, что в этом случае она могла сохраниться под землей. Дело в том, что организмы, живущие в слоях земли, защищены от любых проблем, которые происходят на суше.
Например, даже если Земля улетит в открытый космос и тут настанет кромешный холод, организмы, которые сейчас живут в толще Земли, сохранят свою жизнь. Их будет греть раскаленное земное ядро и магма.
Вот почему жизнь могла остаться и на Марсе.
Впрочем, это только гипотеза. Скорее всего, на Марсе никогда и не было жизни. Ведь наличие воды и атмосферы – это совсем не гарантия зарождения жизни.
Я вообще прагматик , полагаю ,что в солнечной системе ЛЮДИ есть только на планете Земля.
Надо беречь и Землю и Людей .
Отредактировано: Пасечник - 13 июн 2020 06:35:16
Кемерово
Карма: +3.14
Регистрация: 30.09.2014
Сообщений: 1,230
Читатели: 0
Регистрация: 30.09.2014
Сообщений: 1,230
Читатели: 0
В середине ХХ века Фред Хойл высказал в «Чёрном облаке» мысль, что биосинтез стартовал на стадии протопланетного облака. Действительно, в случае протооблака налицо несравненно бОльшие ресурсы как по массе и объёму реагентов, так и по времени. Поэтому если самопроизвольное образование механизма ДНК могло иметь место, то именно ДО образования планет. Соответственно планетное вещество было пропитано той протоорганикой – о чём, например, свидетельствует НЕФТЬ, следы которой обнаружены на Кольской сверхглубокой.
Таким образом, жизнь на планетах системы исключать никак нельзя. А вот разумная жизнь – это исключительно по части Всевышнего.
Таким образом, жизнь на планетах системы исключать никак нельзя. А вот разумная жизнь – это исключительно по части Всевышнего.
Сейчас на ветке:
4,
Модераторов: 0,
Пользователей: 0,
Гостей: 0,
Ботов: 4